|
A neuron has a cell body with extensions leading off it. Numerous dendron and dendrites provide a large surface area for connecting with other neurons, and carry nerve impulses towards the cell body. A single long axon carries the nerve impulse away from the cell body. The axon is only 10µm in diameter but can be up to 4m in length in a large animal (a piece of spaghetti the same shape would be 400m long)! Most neurons have many companion cells called Schwann cells, which wrap their cell membrane around the axon many times in a spiral to form a thick insulating lipid layer called the myelin sheath. Nerve impulse can be passed from the axon of one neuron to the dendron of another at a synapse. A nerve is a discrete bundle of several thousand neuron axons. Humans have three types of neuron:
Motor neurons Interneuron's |
(Hasibah, 2004)
| Neurones and muscle cells are electrically
excitable cells, which means that they can transmit electrical nerve
impulses. These impulses are due to events in the cell membrane, so to
understand the nerve impulse we need to revise some properties of cell
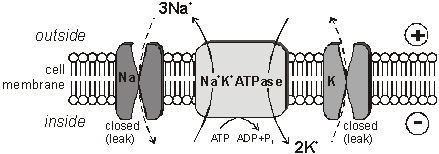
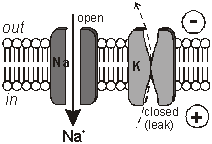
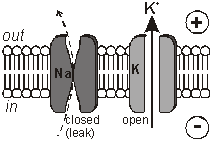
membranes. The Membrane Potential All animal cell membranes contain a protein pump called the Na+K+ATPase. This uses the energy from ATP splitting to simultaneously pump 3 sodium ions out of the cell and 2 potassium ions in. If this was to continue unchecked there would be no sodium or potassium ions left to pump, but there are also sodium and potassium ion channels in the membrane. These channels are normally closed, but even when closed, they "leak", allowing sodium ions to leak in and potassium ions to leak out, down their respective concentration gradients.
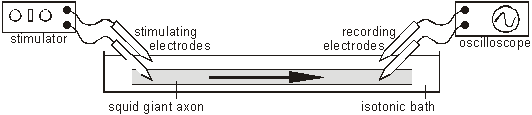
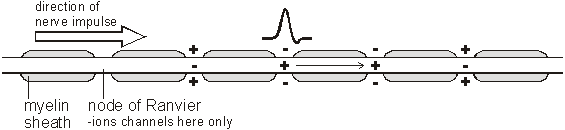
The combination of the Na+K+ATPase pump and the leak channels cause a stable imbalance of Na+ and K+ ions across the membrane. This imbalance causes a potential difference across all animal cell membranes, called the membrane potential. The membrane potential is always negative inside the cell, and varies in size from –20 to –200 mV in different cells and species. The Na+K+ATPase is thought to have evolved as an osmoregulator to keep the internal water potential high and so stop water entering animal cells and bursting them. Plant cells don’t need this as they have strong cells walls to prevent bursting. The Action Potential In nerve and muscle cells the membranes are electrically excitable, which means that they can change their membrane potential, and this is the basis of the nerve impulse. The sodium and potassium channels in these cells are voltage-gated, which means that they can open and close depending on the voltage across the membrane. The nature of the nerve impulse was discovered by Hodgkin, Huxley and Katz in Plymouth in the 1940s, for which work they received a Nobel prize in 1963. They used squid giant neurones, whose axons are almost 1mm in diameter, big enough to insert wire electrodes so that they could measure the potential difference across the cell membrane. In a typical experiment they would apply an electrical pulse at one end of an axon and measure the voltage changes at the other end, using an oscilloscope:
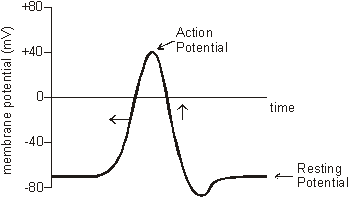
The normal membrane potential of these nerve cells is –70mV (inside the axon), and since this potential can change in nerve cells it is called the resting potential. When a stimulating pulse was applied a brief reversal of the membrane potential, lasting about a millisecond, was recorded. This brief reversal is called the action potential:
The action potential has 2 phases called depolarisation and repolarisation. Depolarisation. The stimulating electrodes cause the membrane potential to change a little. The voltage-gated ion channels can detect this change, and when the potential reaches –30mV the sodium channels open for 0.5ms. The causes sodium ions to rush in, making the inside of the cell more positive. This phase is referred to as a depolarisation since the normal voltage polarity (negative inside) is reversed (becomes positive inside).
Repolarisation. When the membrane potential reaches 0V, the potassium channels open for 0.5ms, causing potassium ions to rush out, making the inside more negative again. Since this restores the original polarity, it is called repolarisation.
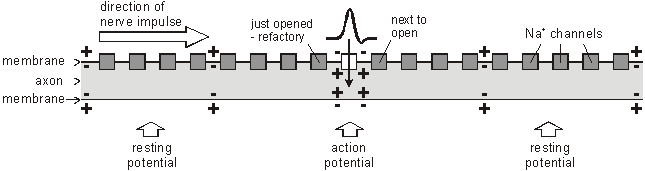
The Na+K+ATPase pump runs continuously, restoring the resting concentrations of sodium and potassium ions. How do Nerve Impulses Start? In the squid experiments the action potential was initiated by the stimulating electrodes. In living cells they are started by receptor cells. These all contain special sodium channels that are not voltage-gated, but instead are gated by the appropriate stimulus (directly or indirectly). For example chemical-gated sodium channels in tongue taste receptor cells open when a certain chemical in food binds to them; mechanically-gated ion channels in the hair cells of the inner ear open when they are distorted by sound vibrations; and so on. In each case the correct stimulus causes the sodium channel to open; which causes sodium ions to flow into the cell; which causes a depolarisation of the membrane potential, which affects the voltage-gated sodium channels nearby and starts an action potential. How are Nerve Impulses Propagated? Once an action potential has started it is moved (propagated) along an axon automatically. The local reversal of the membrane potential is detected by the surrounding voltage-gated ion channels, which open when the potential changes enough.
The ion channels have two other features that help the nerve impulse work effectively:
How Fast are Nerve Impulses? Action potentials can travel along axons at speeds of 0.1-100 m/s. This means that nerve impulses can get from one part of a body to another in a few milliseconds, which allows for fast responses to stimuli. (Impulses are much slower than electrical currents in wires, which travel at close to the speed of light, 3x108 m/s.) The speed is affected by 3 factors:
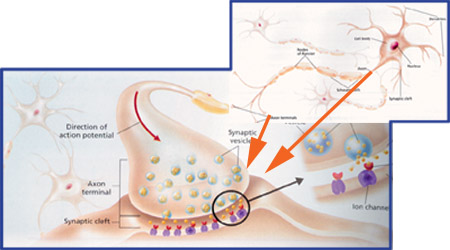
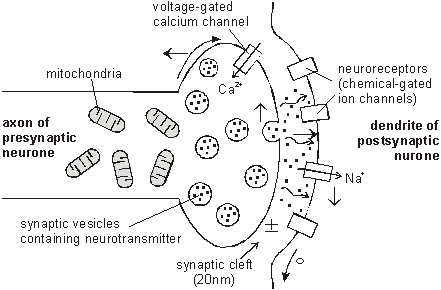
synapse The junction between two neurons is called a synapse. An action potential cannot cross the synaptic cleft between neurons, and instead the nerve impulse is carried by chemicals called neurotransmitters. These chemicals are made by the cell that is sending the impulse (the pre-synaptic neuron) and stored in synaptic vesicles at the end of the axon. The cell that is receiving the nerve impulse (the post-synaptic neuron) has chemical-gated ion channels in its membrane, called neuroreceptors. These have specific binding sites for the neurotransmitters
1. At the end of the pre-synaptic neuron there are voltage-gated calcium channels. When an action potential reaches the synapse these channels open, causing calcium ions to flow into the cell. 2. These calcium ions cause the synaptic vesicles to fuse with the cell membrane, releasing their contents (the neurotransmitter chemicals) by exocytose. 3. The neurotransmitters diffuse across the synaptic cleft. 4. The neurotransmitter binds to the neuroreceptors in the post-synaptic membrane, causing the channels to open. In the example shown these are sodium channels, so sodium ions flow in. 5. This causes a depolarization of the post-synaptic cell membrane, which may initiate an action potential. 6. The neurotransmitter is broken down by a specific enzyme in the synaptic cleft; for example the enzyme acetylcholinesterase breaks down the neurotransmitter acetylcholine. The breakdown products are absorbed by the pre-synaptic neurone by endocytosis and used to re-synthesise more neurotransmitter, using energy from the mitochondria. This stops the synapse being permanently on. Different Types of Synapse The human nervous system uses a number of different neurotransmitter and neuroreceptors, and they don’t all work in the same way. We can group synapses into 5 types: 1. Excitatory Ion Channel Synapses. These synapses have neuroreceptors that are sodium channels. When the channels open, positive ions flow in, causing a local depolarisation and making an action potential more likely. This was the kind of synapse described above. Typical neurotransmitters are acetylcholine, glutamate or aspartate. 2. Inhibitory Ion Channel Synapses. These synapses have neuroreceptors that are chloride channels. When the channels open, negative ions flow in causing a local hyperpolarisation and making an action potential less likely. So with these synapses an impulse in one neurone can inhibit an impulse in the next. Typical neurotransmitters are glycine or GABA. 3. Non Channel Synapses. These synapses have neuroreceptors that are not channels at all, but instead are membrane-bound enzymes. When activated by the neurotransmitter, they catalyse the production of a "messenger chemical" inside the cell, which in turn can affect many aspects of the cell’s metabolism. In particular they can alter the number and sensitivity of the ion channel receptors in the same cell. These synapses are involved in slow and long-lasting responses like learning and memory. Typical neurotransmitters are adrenaline, noradrenaline (NB adrenaline is called epinephrine in America), dopamine, serotonin, endorphin, angiotensin, and acetylcholine. 4. Neuromuscular Junctions. These are the synapses formed between motor neurones and muscle cells. They always use the neurotransmitter acetylcholine, and are always excitatory. We shall look at these when we do muscles. Motor neurones also form specialised synapses with secretory cells. 5. Electrical Synapses. In these synapses the membranes of the two cells actually touch, and they share proteins. This allows the action potential to pass directly from one membrane to the next. They are very fast, but are quite rare, found only in the heart and the eye. Summation
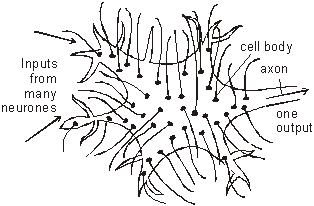
One neuron can have thousands of synapses on its body and dendrons. So it has many inputs, but only one output. The output through the axon is called the Grand Postsynaptic Potential (GPP) and is the sum of all the excitatory and inhibitory potentials from all that cell’s synapses. If there are more excitatory potentials than inhibitory ones then there will be a GPP, and the neuron will "fire", but if there are more inhibitory potentials than excitatory ones then there will not be a GPP and the neuron will not fire. This summation is the basis of the processing power in the nervous system. Neurons (especially interneuron's) are a bit like logic gates in a computer, where the output depends on the state of one or more inputs. By connecting enough logic gates together you can make a computer, and by connecting enough neurons together to can make a nervous system, including a human brain.
|
***************************@@@@@@***************************
Copyright © Hasibah Mohammad - University Of Twente (Netherlands, 2004)
Acknowledgement for
Java Web Start - Java Compiler
Mark Rothery's Biology Web Site http://www.mrothery.co.uk/nerves/NervesNotes.htm